
Grouping and Cooperation
Contents
Grouping and Cooperation¶
There is a spectrum of social behavior that ranges from not solitary to eusocial. The categories along this spectrum are shifty. The main concepts considered today are grouping and cooperation.
Case Study: An Insect’s Extreme Makeover¶

MARTIN ENSERINK, Science 2004
Schistocerca gregaria, the desert locust, is a dull-looking, shy insect that tends to stay put, avoid other locusts, fly by night, and never cause trouble. And then there’s the desert locust, Schistocerca gregaria, a conspicuous yellow-and-black—or bright pink when not fully mature—thrill seeker that bands together in swarms of billions that cross vast distances in broad daylight and devour tons of vegetation in their path.
So striking is the difference between the desert locust’s “solitary” and “gregarious” phases that it wasn’t until 1921 that Russian entomologist Boris Uvarov realized they were the same species. And only recently have scientists begun to piece together a detailed picture of how the insect switches from one phase to the other. University of Oxford entomologist Stephen Simpson, the uncontested leader in this small field, hopes that this understanding may eventually help prevent plagues. “The phase change is the defining feature of locust biology,” he says,“and also the main problem.”
The makeover is the locust’s answer to harsh life in the desert, Simpson explains. Most of the time, the sparse vegetation can sustain only small numbers of desert locusts, and they do best by staying out of one another’s way. After intense rain, however, plant life explodes and locust numbers skyrocket; when the inevitable drought sets in, the insects find themselves coalescing in high numbers around shrinking food supplies. This increased density is what triggers the shift from solitary to gregarious—presumably because, once they run out of food, the insects need to migrate and, like many species, they seek safety in numbers.
Researchers have long known that the locust’s behavior is the first thing to change. A solitary locust becomes more attracted to its mates and more active after spending just 4 hours in a crowded cage, for instance. The spectacular morphological transformation, on the other hand, can take several generations to complete.
When densities drop (for instance, when enough members of a swarm die) the process reverses.
⏳ 10 min
Q1: What is an ultimate explanation provided by Enserink for social behavior in locusts? What is a proximate explanation provided by Enserink for the phenotype switch?
Q2: In general, why might individuals be better off solitary? (ie. what are some negative fitness effects of grouping).
Q3: In general, what functions can you think of for grouping? (ie how could grouping increase individual fitness?)
⏸️ PAUSE here for class-wide discussion
Why group?¶
Passive Defense¶
When the only anti-predator “action” taken is to group.
Case Study: Dilution Effect¶
Grouping may reduce the probability of any one individual being captured by predators.
Burger et al1 tested this effect in butterflies that “puddle” together in groups of various sizes to drink water.
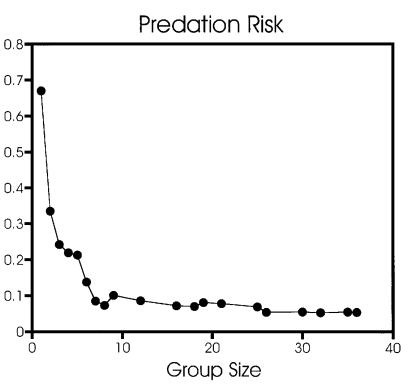
Fig. 123 Risk to individual pierid butterflies of capture by smooth-billed anis as a function of the number of butterflies in puddle-drinking clusters. For each cluster size, the risk was calculated by dividing the mean number of predator-captured butterflies by the cluster size. The number of butterflies captured increased as the total number of butterflies increased, but reached a threshold beyond which the capture rate did not increase. An individual ani was unable to catch more than 16, regardless of group size. From Burger and Gochfeld 20011¶
Case Study: Confusion Effect¶
Grouping may reduce the likelihood of predators capturing prey because of their inability to single out and attack individual prey. It confuses predators that have difficulty tracking a single object.
Hogan et al2 used a video game simulation to test this hypothesis. The experimental setting resembled that of a flight simulator. A human “predator” used a game controller to steer an avian predator through a virtual three-dimensional environment. Participants were presented with a crosshair over a randomly selected bird in the simulated starling flock and were tasked with steering toward and “killing” the targetted bird.
Active Defense¶
Specicalized anti-predator behaviors beyond grouping.
Case Study: The effect of group size on vigilance¶

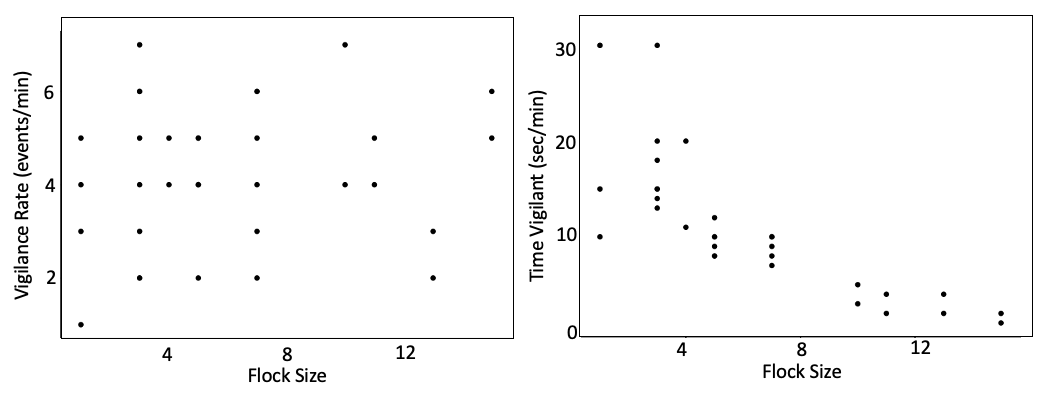
Fig. 126 Example results from analysis of vigilance behavior from this video. Time vigilant varied with flock size. Vigilance rate was not an effective metric at capturing this phenomenon.¶
⏳ 5 min
Q4: Explain a potential function for grouping. Formulate the explanation based both on these results and on the conflict between feeding and vigilance behaviors.
⏸️ PAUSE here for class-wide discussion
Case Study: Mobbing¶
Black-headed gulls are a colonial ground-nesting species that mob potential predators (as a group: dive-bombing predators, pooping on them, etc). Mobbing takes time and energy, increases the likelihood of injury or death, and such a gregarious display can end up attracting more predators. So why does mobbing behavior exist (and why in this species)?
One hypothesis is that that mobbing potential predators distracts them from finding and therefore predating gull egg nests.
One prediction of this anti-predator hypothesis is that mobbing should force distracted predators to expend more searching effort than they would otherwise. Kruuk (1964) observed and described that mobbed crows could not look around “comfortably” for gull nests when they are being mobbed, and therefore are probably less likely to find them without increasing their effort.
Another prediction is that the benefit experienced by mobbing gulls should be directly proportional to the extent that predators are actually mobbed (ie. if the function if mobbing is to reduce egg predation, then mobbing should reduce egg predation). Kruuk also tested this by placing chicken eggs along a line that spanned from the inside to the outside a gull nesting colony.
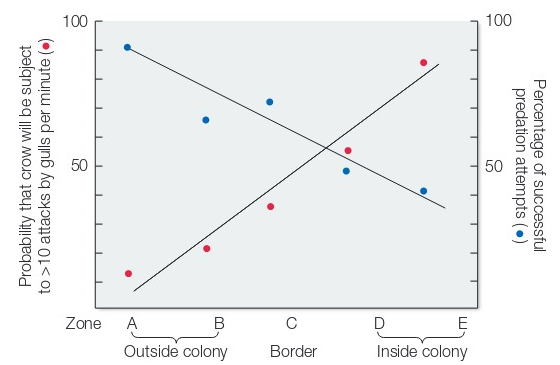
Fig. 127 Crows were attacked more inside the gull colony (red). Crows ate less of the eggs inside the gull colony (blue).¶
We can also implement a comparative (ie phylogenetic and evolutionary) approach to explain mobbing in gulls. If mobbing is “an evolved response to predation on eggs and nestlings,” then a key prediction is that gull species whose nests were at a lower risk of predation should not exhibit mobbing behavior. In other words, if the risk of nest predation is lower, then there is less of a positive selective pressure on mobbing (the “reduces predation” factor” of the evolutionary model affects fitness less). This would alter the oucome of an evolutionary cost-benefit model.
Kittiwakes do not mob their predators. Kittiwakes are an example of a gull species that nest on vertical coastal cliffs. Cliff-nesting species have relatively few nest predators because it is hard for small mammals to scale cliffs in search of prey and predatory birds have difficulty maneuvernig near cliffs in turbulent coastal wind. Additionally, kittiwake adults are smaller than black-headed gulls and therefore more vulnerable to predator attacks themselves (ie. this negative factor would affect fitness more).
Therefore, a comparative analyiss of black-headed gulls and kittiwakes seem to support this key evolutionary prediction of the anti-predator hypothesis.

Additionally, distantly-related colonial, ground nesting birds (like bank swallows) have evolved mobbing behavior. Even some colonially nesting mammals have evolved mobbing behavior (for example California ground squirrels Otospermophilus beecheyi).
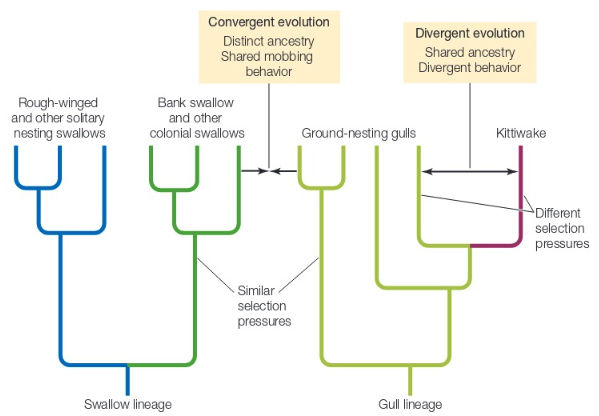
Fig. 128 Convergent and divergent evolution of mobbing behavior across swallows (Hirundinidae) and gulls (Laridae). The last common ancestor of swallows and gulls occurred long ago and they are in different families. All else being equal, such distantly related animals would be expected to behave differently (especially if their last common ancestor did not share the behavioral trait). More similarly related species (for example within Laridae) would be expected to behave similarly because of how much genetic material they share.¶
Altruism¶
Mutual benefit is when a social behavior results in increased fitness of all individuals involved (like mobbing). Altruism occurs when the recipient of a social act gains fitness, but the donor of the social act decreases its immediate fitness. Therefore, the existence of altruistic behavior presents a paradox for evolutionary theorists because fitness is thought to be the main variable effecting evolutionary change (ie natural selection).
At first, it may seem that cooperative altruism could result in mutual benefit when averaged over time (ie. when everyone you encounter are also altruists, you lose fitness by helping others, but you gain fitness when others help you). However, selfishness is the evolutionary thorn in the side of altruism. Selfishness occurs when an individual does not reciprocate an altruistic act.
An evolutionary examination of altruism is often formulated in a game theory model in which:
When altruists interact, they exchange the cost and benefit of altruism
When altruists interact with selfish individuals, the altruist pays the cost and the selfish individual receives the benefit
When selfish individuals interact, they receive no benefit but expend no cost.
The basic payoff structure is shown below:
Altruist |
Selfish |
|
|---|---|---|
Altruist |
(b - c) / 2 |
-c |
Selfish |
b |
0 |
⏳ 5 min
Q5: Using this game theory model, show why you would never predict to observe altruistic cooperation.
⏸️ PAUSE here for class-wide discussion
Case Study: Migration in Pelicans and Ibis¶

Fig. 129 Catching air. Northern bald ibises (Geronticus eremita) migrate in the perfect V-formation. MARKUS UNSÖLD (WALDRAPPTEAM).4¶
These aerodynamic accomplishments were previously not thought possible for birds because of the complex flight dynamics and sensory feedback that would be required to perform such a feat5.
We now also know that birds at different positions in the formation explicitly expend different amounts of energy.
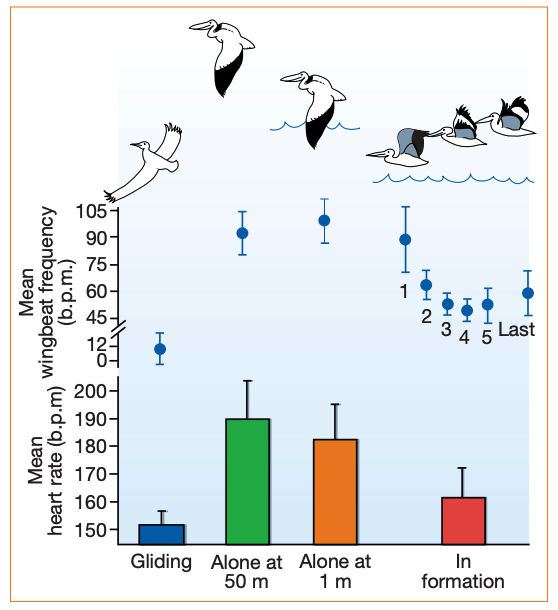
Fig. 130 Wing-beat frequency and heart rate of pelicans engaged in various types of flight (mean ± s.d.)[^Weimerskirch-2001].¶
But why does any individual bird fly at the front? At the front there is no aerodynamic advantage and, unlike in solo flight, they flap continuously without gliding (presumable to increase the aerodynamic boost to the trailing birds).
We do know6 that Northern bald ibis (Geronticus eremita) match the time they spend in the wake of each other by frequent pairwise switches of the leading position (ie. evidence for cooperative altruism).
Given that humans and other animals do cooperate, the challenge to the evolutionary theorist has been to find some mechanism that will permit altruism to resist elimination by selfish alternatives.
Some argue that the payoff structure of in-wake flying resembles the structure of a snowdrift model – one among a variety of proposed solutions to the cooperation dilemma.
The Snowdrift model derives its name from the following situation: two drivers are trapped on either side of a snowdrift and have the options of staying in the car or removing the snowdrift. Letting the opponent do all the work is the best option. But if the other player stays in the car, it is better to shovel because you still obtain a benefit from your behavior in addition to the cost (Sugden 1986). In this game, cooperation yields an increase in fitness (b) that is accessible to both players (i.e. free passage to go home), whereas the decrease to fitness (c; i.e. removing the snowdrift) is shared between cooperators.
The basic payoff structure is shown below (values in parenthesis are an example under the conditions b > c):
Cooperator (Altruist) |
Selfish |
|
|---|---|---|
Cooperator (Altruist) |
b - (c/2) (200) |
b - c (100) |
Selfish |
b (300) |
0 (0) |
In the snowdrift game, a noncooperator meeting a cooperator gains the highest payoff, but a noncooperator meeting another noncooperator (“selfish in selfish population’) can earn even less from an interaction than a cooperator meeting a noncooperator (“altruist mutant in selfish population”) – leading to the possibility of a mixed phenotype population. This result is in contrast to the prediction of the basic altruist/selfish model (which predicts that the selfish phenotype is the only evolutionarily stable solution).
Green Beard¶
Additional Resources¶
Weimerskirch, H., Martin, J., Clerquin, Y. et al. Energy saving in flight formation. Nature 413, 697–698 (2001).
Portugal, S., Hubel, T., Fritz, J. et al. Upwash exploitation and downwash avoidance by flap phasing in ibis formation flight. Nature 505, 399–402 (2014).
Raw Data from behavioR package with the following script to convert to csv:
!pip install rdata import rdata import pandas as pd path = '/content/MeerkatScanData.rda' parsed = rdata.parser.parse_file(path) converted = rdata.conversion.convert(parsed) converted['MeerkatScanData'].to_csv('MeerkatScanData.csv')Serotonin Mediates Behavioral Gregarization Underlying Swarm Formation in Desert Locusts
Phenotypic Transformation Affects Associative Learning in the Desert Locust
- 1(1,2)
- 2(1,2)
The confusion effect when attacking simulated three-dimensional starling flocks
- 3
- 4
[Why Birds Fly in a V Formation: Study in ibises suggests the animals are giving each other a lift (2014)]https://www.science.org/content/article/why-birds-fly-v-formation
- 5
- 6